Меню сайта
- На главную
- Рост растений
- Почвенные простейшие
- Полиморфизмы
- Дыхание бактерий
- Органическая природа
- Первая жизньу
- Материалы по биологии
Последние новости
Причина отравления вод океана.
Американские ученые из штата Мичиган полагают, что в качестве главной причины отравления вод Мирового океана ртутью являются бактерии.
Секрет выживания лягушек.
Американским ученым удалось выяснить, как лягушкам удается продолжать жить даже после глубокой заморозки.
Секрет долголетия ночницы.
Биологи уже давно считают, что продолжительность жизни животного определяется очень просто: чем оно больше, тем дольше живет.
Выбор физической модели кинетики энергетического метаболизмаМатериалы по биологии и химии / Исследование изменений метаболизма эритроцитов донорской крови после взятия ее из организма / Выбор физической модели кинетики энергетического метаболизмаСтраница 1
Метаболизм большинства соматических клеток, выполняющих определенные функции в организме, описывается огромным количеством параметров, что, как правило, существенным образом затрудняет создание математической модели их состояний. Исключение составляют только отдельные виды клеток с простейшим метаболизмом, к которым относится эритроцит. Однако физические модели, основанные на качественном анализе закономерностей протекания конкретных процессов метаболизма, могут дать значимые результаты для решения фундаментальных и прикладных задач. В данной работе использовались только физические модели, которые более просты и наглядны. Блефаропластика верхних и нижних нижняя блефаропластика.
В литературе наиболее известна модель Селькова [14], используемая в работе Атауллаханова. Математическая модель сложного равновесного процесса усвоения глюкозы по пути Эмбдена-Мейэргофа представлена системой уравнений, описывающих каждую реакцию последовательного превращения молекулы глюкозы в лактат. Расчеты приводятся при постоянном значении рН и при постоянстве пула ферментов и регуляторных субстанций, участвующих в этих реакциях. В результате получают зависимость скоростей утилизации глюкозы от концентрации АТФ в цитоплазме эритроцита, то есть зависимость скорости V+ синтеза АТФ от его концентрации nАТФ. В модели принято, что расход АТФ обусловлен суммарной тратой этого макроэрга в простых ферментативных реакциях. Следовательно, скорость V- трат АТФ в зависимости от nАТФ в цитоплазме эритроцита изменяется по закону Михаэлиса-Ментен:
![]() ,
,
где ![]() - скорость траты АТФ,
- скорость траты АТФ, ![]() - концентрация АТФ, величины km и А - константы. В данной модели km>> nАТФ, тогда
- концентрация АТФ, величины km и А - константы. В данной модели km>> nАТФ, тогда
![]()
Данная модель представлена на рис.3.
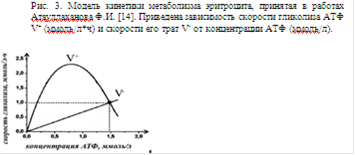
При пересечении кривых V+ и V- реализуется только одна стационарная точка. Устойчивость точки определяется по двум условиям устойчивости по Ляпунову:
(1) - первое условие устойчивости,
где αi - рассматриваемый параметр, ![]() и
и ![]() -соответственно, скорости притока и убыли данного параметра, αj - параметр, по которому проверяется устойчивость, при этом i может быть равно j.
-соответственно, скорости притока и убыли данного параметра, αj - параметр, по которому проверяется устойчивость, при этом i может быть равно j.
(2) - второе условие устойчивости.
![]()
Стационарная точка является устойчивой по условию (1). Видно, что данная модель не допускает существования большего количества устойчивых стационарных точек.
Следовательно, можно заключить, что для установления возможности существования устойчивых точек с помощью моделей, в которых допускается свободной одна переменная, достаточно знать вид кривых зависимостей скоростей синтеза V+ и расхода V- АТФ от его концентрации и их взаимное расположение. Такой графический метод анализа кинетики метаболизма наиболее нагляден и позволяет на качественном уровне устанавливать наличие устойчивых стационарных точек, поэтому в дальнейшем будет решаться поставленная задача данным методом, без привлечения сложных математических уравнений.
Другой физической моделью, схожей с предыдущей, является модель, приведенная в работах Фока М.В. и его соавторов [10]. Она используется для анализа кинетики энергетического метаболизма и представлена на рис. 4. Данная модель принята за основную в данном исследовании.

Зависимости скоростей синтеза (V+) и расхода (V-) АТФ от его концентрации (nАТФ) в цитоплазме при постоянстве всех остальных параметров, таких как рН, концентраций всех ферментов, количества аденозиновых оснований (АДФ и АМФ) и запасов неорганического фосфата, имеют вид, представленный на рис. 4. Свободным параметром, также как и в первой модели, является концентрация АТФ в цитоплазме.
Кривая зависимости V+ от nАТФ имеет колоколообразную форму с максимумом. При нулевой концентрации АТФ в цитоплазме скорость синтеза V+ АТФ имеет ненулевое значение. Это обусловлено тем, что в рассматриваемых условиях (заданном значении рН, концентрации АДФ, ограниченном запасе неорганического фосфата и при нулевом значении концентрации АТФ) в результате теплового движения субстраты (АДФ и молекул фосфата) сталкиваются, вследствие чего преодолевается активационный барьер, и в результате, происходит синтез АТФ. Дальнейший ход кривой зависимости скорости (V+) синтеза АТФ от nАТФ определяется автокаталитической реакцией синтеза, в которой часть выработанного АТФ расходуется в процессах гликолитических реакций. Поэтому скорость синтеза АТФ сначала зависит линейно от nАТФ, а затем экспоненциально. Далее по мере истощения запасов органического фосфата достигается максимум скорости синтеза АТФ. Дальнейший спад кривой зависимости V+ АТФ от nАТФ обусловлен, в основном, истощением запасов неорганического фосфата. При нулевой скорости синтеза в области высоких значений nАТФ все запасы неорганического фосфата израсходованы.
Смотрите также
Ботанические сады Украины
Введение
В своем реферате я рассмотрю один из главных туристических ресурсов,
который всегда пользовался большой популярностью среди туристов. Это
ботанический сад. Только здесь можно ...
Влияние ауксина на рост и развитие растений
...
Возбудитель стахиботриотоксикоза
Введение
Проблема
микотоксинов и микотоксикозов животных в настоящее время стала не только
экономически, но и социально значимой. Были произведены исследования по
выявлению грибов, о ...